Teleoceras proterum

Quick Facts
Common Name: Archer short-legged rhino
The mounted skeleton on display at the Florida Museum of Natural History (see image) was constructed as part of a “live” public display during the late 1970s using fossils from both the McGehee Farm and Love Bone Bed localities.
Thousands of fossils of Teleoceras proterum have been found at two different sites in Florida, suggesting that it was a very common species during the time it was living.
Despite its similarities in body plan to the modern hippopotamus (short limbs, large barrel-shaped rib cage), the majority of scientific evidence favors a terrestrial way of life over an amphibous, hippo-like ecology.
Age Range
- Late Miocene epoch; late Clarendonian (Cl3) to early Hemphilian (Hh1) land mammal ages
- About 9.5 to 8 million years ago
Scientific Name and Classification
Teleoceras proterum Leidy, 1885
Source of Species Name: Not explicitly stated in Leidy’s (1885:33) original description. The species name may have been formed from the Greek word proteros meaning ‘earlier’.
Classification: Mammalia, Eutheria, Laurasiatheria, Perissodactyla, Ceratomorpha, Rhinocerotoidea, Rhinocerotidea, Teleoceratini
Alternate Scientific Names: Rhinoceros proterus, Aphelops fossiger, Teleoceras fossiger
Overall Geographic Range
This species is endemic to Florida. The type locality is Mixsons Bone Bed, Levy County, Florida (Leidy, 1885; Prothero, 2005).
Florida Fossil Occurrences
Florida fossil sites with Teleocera proterum:
- Alachua County—Haile 19A; Love Site; McGehee Farm
- Levy County—Mixson’s Bone Bed
- Polk County—Four Corners Fauna (Amalgamated Phosphate Company Mine; Palmetto Mine)
Discussion
Perissodactyla is an order that includes mammals like horses (Equidae), tapirs (Tapiridae), and rhinoceroses (Rhinoceratidae). While extant rhinoceroses are only found in Africa and Asia, they were also widely distributed in North America and Europe in the past (Prothero, 2005). Teleoceras proterum is an extinct species of rhinoceros that was present in the late Miocene of Florida.
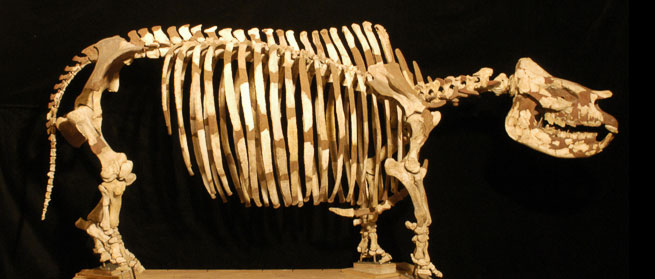
Fossils of Teleoceras proterum were first found at Mixson’s Bone Bed locality in Levy County, Florida, where several thousands of specimens representing a minimum of 117 individuals were discoverd (Prothero, 2005). The Mixson’s sample is divided between the National Museum of Natural History in Washington, the American Museum of Natural History in New York (which has the largest collection), and the Florida Museum of Natural History. The only mounted skeleton of the species is on permanent display at the Florida Museum of Natural History (Fig. 2).
The second largest sample of Teleoceras proterum was found at the Love Bone Bed site near Archer, Florida. Almost 2,000 specimens representing a minimum of 42 individuals were recovered there (MacFadden and Hulbert, 1990). Using measurements of the humeri and femora from the Love Bone Bed sample, MacFadden and Hulbert (1990) produced an estimated average body mass of 615 kg (1350 lbs) for this species.
Historically, Teleoceras proterum was known by some alternate names and sometimes (Leidy and Lucas, 1896; Osborn, 1904; Harrison and Manning, 1983) considered to be a subspecies of Teleoceras fossiger, a contemporaneous species found in Nebraska, South Dakota, Kansas, Oklahoma, Texas, and Nevada (Prothero, 2005). However, Teleoceras fossiger is distinguished by being the largest member of Teleoceras while Teleoceras proterum is smaller and does not overlap with the larger species in size. Teleocerus proterum is also on average smaller than Teloceras major, Teleoceras hicksi, and an undescribed late Hemphillian species of Teloceras from Tennessee and Florida (Short, 2013). Based on this size difference, the lack of geographic overlap, and its relatively shorter and more robust limb bones, Teleoceras proterum is currently considered to be a valid species (Prothero, 2005). However, it is likely that these species were all closely related to one another.
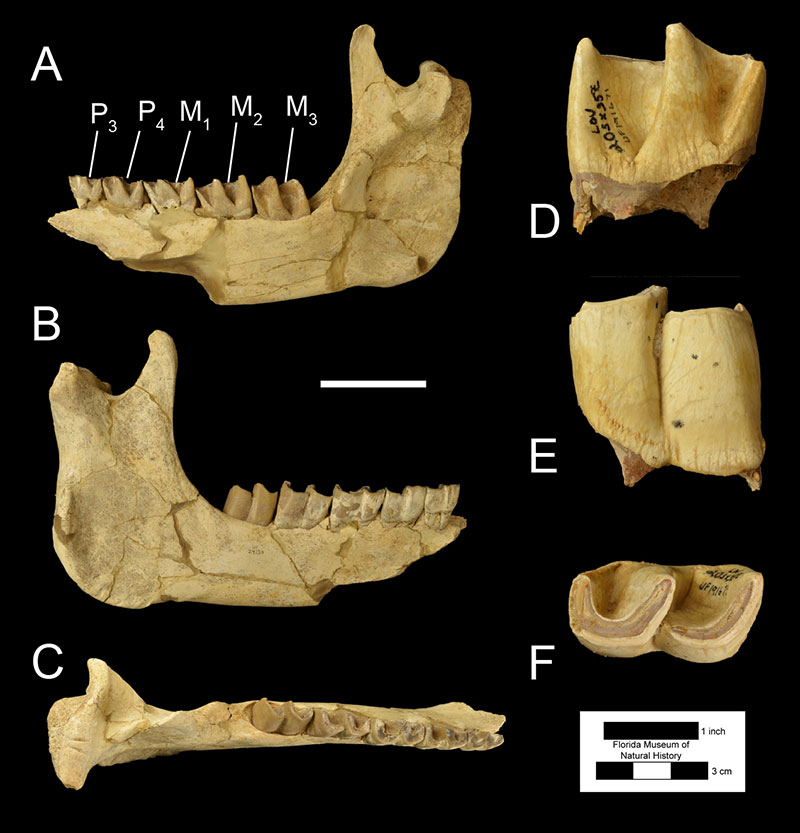
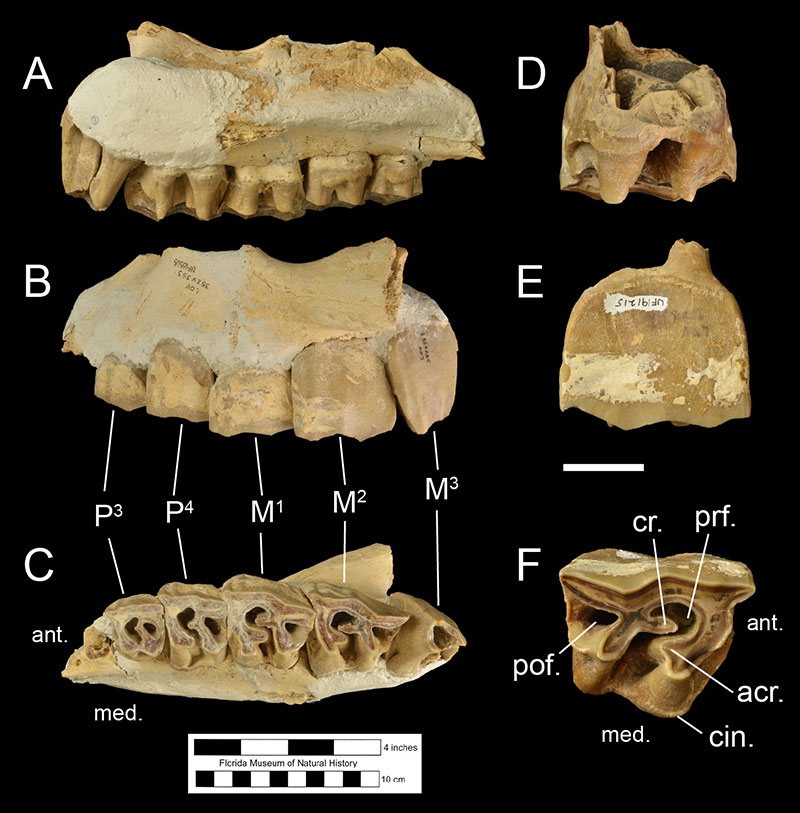
Fossils of Teleoceras proterum may be identified from Florida fossil assemblages using several diagnostic features. Firstly, the teeth of rhinoceroses are distinctive from those of other mammals in their larger size and form. Lower molars are characterized by 2 consecutive L-shaped lophs (Fig. 3), and upper molars are rectangular in outline with features known as fossettes, crochets, and antecrochets (Fig. 4). However, in rhinoceroses of the family Rhinocerotidae (like Teleoceras proterum), the upper third molar is triangular in outline (Fig. 4C; Hulbert, 2001). Furthermore, the molar teeth of Teleoceras proterum can be distinguished from those of other Miocene rhinocerotids found in Florida such as Aphelops and Peraceras by the greater complexity and higher crown of their teeth; Teleoceras proterum had relatively more hypsodont dentition (Hulbert, 2001). Within the genus Teleoceras, Teleoceras proterum is distinguished by having more a more prominent cingulum on the upper third and fourth premolars and by having narrower, shorter crochets and antecrochets on their upper molars (Prothero, 2005). Male and female Teleoceras proterum are sexually dimorphic, with males possessing larger lower second incisors, or tusks (Figure 5). The size of these tusks allows for differentiating between males and females of this species (Mihlbachler, 2003, 2005). Cranially, Teleoceras proterum is similar to other species of Teleoceras in having a relatively brachycephalic (shortened) skull relative to other genera of rhinoceroses (Prothero, 2005). A single, broad upper incisor wears against the lower tusk keeping it sharp, a feature absent in Aphelops.
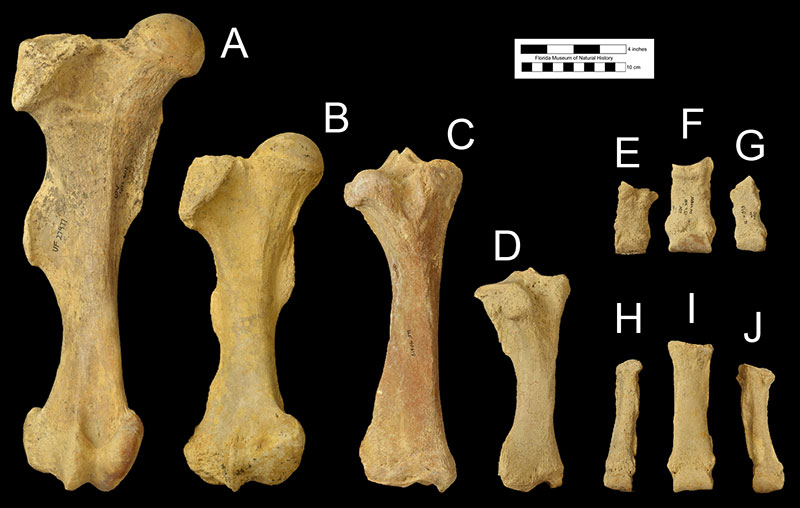
Postcranially, Teleoceras proterum was characterized by a body shape similar to other members of the genus. Much like the hippopotamus (which is not closely related to rhinoceroses, but is more closely related to the cows, deers, and camels), Teleoceras proterum had a very wide, barrel-shaped torso and relatively short, stumpy limbs. This differs greatly from other Miocene Florida rhinocerotids including Floridaceras, Aphelops, or Peraceras, which were relatively long-limbed (Figs. 6 and 7; Hulbert, 2001; Prothero, 2005). In addition, the foot bones of Teleoceras proterum are wider laterally and shorter in the proximal to distal dimension compared to Aphelops, making their differentiation possible in sites where they co-occur (Fig. 7; Harrison and Manning, 1983). Furthermore, among species of its genus, Teleoceras proterum appears to have had the shortest and most robust limb bones (Prothero, 2005).

There is some debate as to whether Teleoceras proterum was semi-aquatic and exhibited social behaviors like the hippopotamus, which is often used as a modern analogue for this fossil species. There is little debate that Teleoceras proterum grazed grasses like hippopotamuses based on the height of their tooth crowns and the wideness of their rostrum, but it is uncertain whether species of Teleoceras spent most of their time in water and came out to graze like hippopotamuses or were primarily terrestrial, as in all extant rhinoceroses (Prothero, 2005). One isotope study of the teeth of Teleoceras proterum (MacFadden, 1998) found that the levels of oxygen isotopes did not suggest that this species was primarily aquatic. On the other hand, further study resulted in some evidence that the isotopes in the original sample may have been contaminated and that in samples of Teleoceras outside of Florida, the oxygen isotopes suggest that this group of rhinoceroses was mostly aquatic in habit (Prothero, 2005).
There has also been much speculation about the degree of similarity between Teleoceras and hippopotamuses in social behavior. In modern hippopotamus populations, females typically form a large group that rest in water and feed together on land, while most males roam as bachelors, for only the dominant males occupy territory in which the females reside (Mihlbachler, 2003). Furthermore, hippopotamuses are not sexually dimorphic. This differs from extant rhinoceroses, which are sexually dimorphic, typically solitary, and have a high level of young male mortality due to combat using their horns and tusks (Mihlbachler, 2005). Thus, if many more fossils of female Teleoceras are found than males at a given locality, it could be argued that A) bachelor males died elsewhere due to a social pattern similar to modern hippopotamuses or B) that there were fewer males in the population due to high level of male death attributed to inter-male competition, much like modern rhinoceroses. While the higher proportion of female Teleoceras major near water at the mass death assemblage at the Ashfall Fossil Bed State Park in Nebraska has been used to support the hippopotamus-like sociality of Teleoceras, Mihlbachler (2003, 2005) argued that based on the levels of sexual dimorphism and patterns of mortality present in Florida Teleoceras proterum populations, there is no specific support that Teleoceras behaved like modern hippopotamuses. In fact, Mihlbachler (2005) concluded that these patterns were more reminiscent of those of modern rhinoceroses.
Sources
- Original Authors: Arianna Harrington, Richard C. Hulbert Jr.
- Original Completion Date: May 28, 2015
- Editor’s Name: Richard C. Hulbert Jr. and Natali Valdes
- Last Updated On: May 30, 2015
This material is based upon work supported by the National Science Foundation under Grant Number CSBR 1203222, Jonathan Bloch, Principal Investigator. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
Copyright © Florida Museum of Natural History, University of Florida