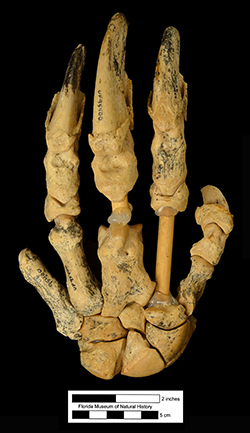
Defining taxon: first appearance of the ground sloth Pliometanastes (Tedford et al., 2004)
Basis of name: Wood et al. (1941) based the name on what they termed the “Hemphill member of the Ogallala [Formation].” This stratigraphic unit was originally named the “Hemphill Beds” by Reed and Longnecker (1932), but is no longer an officially used stratigraphic unit (Gustavson et al., 1990). The stratigraphic name is based on Hemphill County, located in the Texas Panhandle just west of the Oklahoma border. It was chosen as the name for this land mammal age because of the Coffee Ranch vertebrate fossil quarry (also called the Miami locality), a large and well-studied vertebrate fauna located in the southwestern part of this county (Matthew and Stirton, 1930; Dalquest, 1983; Schultz, 1990). The correct pronunciation is ‘hemp-hill-i-an’ and not ‘hem-fill-i-an.’ The standard two-letter abbreviation for the Hemphillian is Hh, as He is used for the Hemingfordian NALMA.
The Hemphillian NALMA is especially well represented by numerous individual vertebrate fossil sites in the central United States (north Texas, Oklahoma, Kansas, eastern Colorado, and Nebraska). The Texas-Oklahoma sites are particularly important, not just for their large numbers of specimens and the diversity and richness of their faunas, but because some are found in stratigraphic superposition above those producing Clarendonian faunas and/or below those producing Blancan faunas, and there are datable ash beds in the region suitable for radioisotopic dating. There are also Hemphillian sites scattered through the western United States and Mexico, and some of these are also amenable to radioisotopic or paleomagnetic dating methods. There are over 200 Hemphillian fossil sites in Florida ranging from Nassau County in north Florida to Lee and Okeechobee counties in south Florida. But most of the Hemphillian sites in Florida are found in two regions of the state. One is in the north-central peninsula, in western Alachua and Marion counties, eastern Gilchrist and Levy counties, and northeastern Citrus County. All but one of the localities in this region are from the early Hemphillian. The second concentration is centered on the “four corners” region of Polk, Hillsborough, Manatee and Hardee counties in south-central Florida. Most of the sites in this region are from the late Hemphillian, but a small number of early Hemphillian sites are known. Other than Florida, elsewhere in eastern North America there are only three major Hemphillian localities, the Mauvilla site in southern Alabama (Hulbert and Whitmore, 2006), the Gray Fossil Site in eastern Tennessee (Hulbert et al., 2009), and the Lee Creek (=Aurora) Mine in eastern North Carolina (Eschelman and Whitmore, 2008). A possible fourth eastern Hemphillian site is the Pipe Creek Sinkhole in Indiana, but there is debate about whether it is latest Hemphillian or earliest Blancan (Farlow et al., 2001; 2010).
The Hemphillian is divided into four subintervals: the Hh1 from 9.0 to 7.5 million years ago; the Hh2 from 7.5 to 6.8 million years ago; the Hh3 from 6.8 to 5.7 million years ago, and the Hh4 from 5.7 to 4.75 million years ago (Tedford et al., 2004). The Hh1 and Hh2 together form the early Hemphillian and the Hh3 and Hh4 the late Hemphillian. There are no fossil localities in Florida that are known to fall in the Hh3 interval.
Index species for Hh1 in Florida: Siren simpsoni, Pseudemys williamsi, Heterodon brevis, Anhinga grandis, Thinobadistes segnis, Mylagaulus kinseyi, Borophagus secundus, Amebelodon floridanus, Prosthennops serus, and Calippus hondurensis
Significant Hh1 localities and faunas in Florida: Haile 19A; McGehee Farm; Tyner Farm; Mixson’s Bone Bed; Gainesville Creeks Fauna; Four Corners Fauna
Index species for Hh2 in Florida: Thinobadistes wetzeli, Borophagus orc, Indarctos oregonensis, Pediomeryx hemphilliensis, and Amebelodon britti.
Significant Hh2 localities and faunas in Florida: Withlacoochee River 4A; Moss Acres Racetrack; Crystal Springs Mobil Home Park; Manatee County Dam; Lockwood Meadows
Index species for Hh4 in Florida: Trachemys inflata, Phoenicopterus floridanus, Ardea polkensis, Alca grandis, Megalonyx curvidens, Borophagus hilli, Eucyon davisi, Agriotherium schneideri, Arctonasua eurybates, Catagonus brachydontus, Mylohyus elmorei, Hemiauchenia edensis, Kyptoceras amatorum, “Dinohippus” mexicanus, Pseudhipparion simpsoni, and Rhynchotherium edense.
Significant Hh4 localities and faunas in Florida: Palmetto Fauna; TRO Quarry; Whidden Creek; Palmetto Mine Microsite; Montbrook; St. Marys River
Characteristic species for the early Hemphillian in Florida: Pseudemys caelata, Macrochelys auffenbergi, Thecachampsa americana, Pliometanastes protistus, Epicyon haydeni, Aepycamelus major, Hemiauchenia minima, Floridameryx floridanus, Metaxytherium floridanum, Pomatodelphis inaequalis, Calippus cerasinus, Calippus maccartyi, Cormohipparion ingenuum, Cormohipparion plicatile, Dinohippus spectans, Nannippus morgani, Nannippus westoni, Neohipparion trampasense, Protohippus gidleyi, Pseudhipparion skinneri, Tapirus webbi, Aphelops malacorhinus, Aphelops mutilus, and Teleoceras proterum.
Characteristic species for the late Hemphillian in Florida: Carcharocles megalodon, Carcharodon hastalis, Galeocerdo cuvier, Hesperotestudo hayi, Borophagus pugnator, Enhydritherium terraenovae, Lynx rexroadensis, Pleiolama vera, Hexameryx simpsoni, Eocoileus gentryorum, Balaenoptera cortesii, Goniodelphis hudsoni, Corystosiren varguezi, Cormohipparion emsliei, Nannippus aztecus, Neohipparion eurystyle, Tapirus polkensis, and Teleoceras hicksi.
Gallery of Hemphillian Florida Fossils
 Carcharocles megalodon
Carcharocles megalodon- Nannippus aztecus
- Megalonyx curvidens
- Eocoileus gentryorum
- Trachemy inflata
- Alca grandis
- Indarctos oregonensis
- Amebelodon floridanus
- Cormohipparion emsliei
- Calippus elachistus
- Aphelops malacorhinus
- Amphimachairodus coloradensis
- Floridameryx floridanus
- Thinobadistes wetzeli
- Aepycamelus major
- Amebelodon britti
- Epicyon haydeni
- Agriotherium schneideri
- Borophagus pugnator
- cf. Alligator mississippiensis (= Alligator meffordi)
- "Dinohippus" mexicanus
- Anhinga grandis





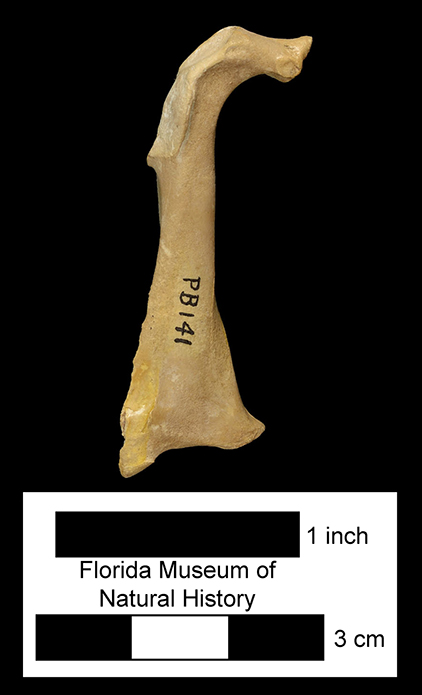




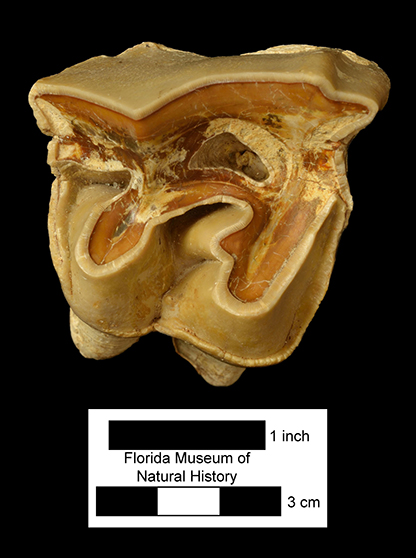










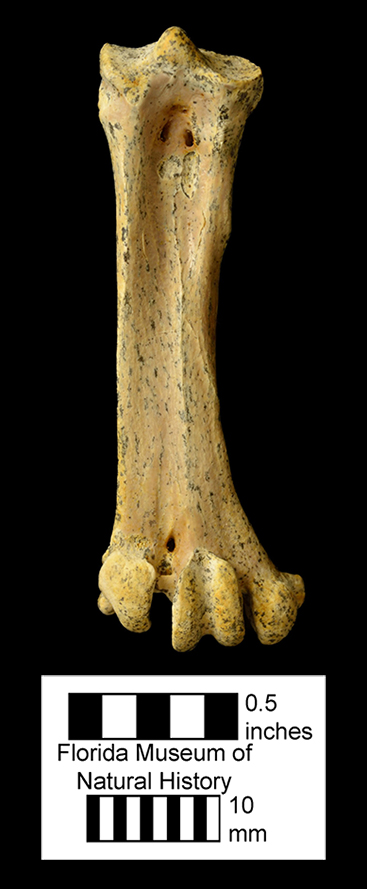
Major extinctions: There are two intervals of wide-spread extinction during the Hemphillian, one at the end of Hh2 and the other at the end of Hh4. The former, the mid-Hemphillian extinction, is widely viewed as being caused by climate change (increased aridity) that lead to a general decrease in forested and other closed habitats and an increase in grasslands and savannas. Browsers and mixed-feeders were more severely impacted than were grazers, that generally became more abundant. Genera that became extinct at this time are the sloth Thinobadistes, the canid Epicyon, the barbourofelid Barbourofelis, the felid Nimravides, the peccary Prosthennops, the camelids Aepycamelus and Procamelus, the protoceratid Synthetoceras, the horses Calippus, Protohippus, Pliohippus, and “Hipparion”, and the gomphothere Amebelodon. There are no definitive records of the rhino Aphelops in the late Hemphillian of Florida, but the genus survived into that interval in western North America. Genera that became extinct during the terminal Hemphillian extinction event include the dugong Nanosiren, the omnivorous canid Carpocyon, the procyonid Arctonasua, the saber-toothed felids Amphimachairodus and Rhizosmilodon, the llama Pleiolama, the camel Megatylopus, the protoceratid Kyptoceras, the pronghorns Hexameryx and Subantilocapra, the deer Eocoileus, the gelocid Floridameryx, the horses Astrohippus, Neohipparion, and Pseudhipparion, and the gomphothere Gomphotherium. The rhino Teleoceras is usually included in this list, but it apparently survived, at least locally, into the earliest Blancan (Gustafson, 2012).
Comments: As was the case elsewhere in the United States, the fauna of the Hh1 interval in Florida is very similar to that of the late Clarendonian (Cl3), and it can be difficult to distinguish Cl3 and Hh1 localities with a limited number of species. Those from the Hh1 will contain either or both of the sloths Thinobadistes segnis and Pliometanastes protistus, the shovel-tusked gomphothere Amebelodon floridanus, the gelocid Floridameryx floridanus, and the peccaries Mylohyus longirostris and Prosthennops serus. The larger species of Calippus from the Cl3, Calippus cerasinus, is replaced by Calippus hondurensis in the Hh1. But most of the common species of horses, rhinos, tapir, and camels are shared between the Cl3 and the Hh1.
The Hh2 interval in Florida shows broad continuity with the Hh1, especially at the genus-level, but in many cases there is a difference at the species-level. For example, Neohipparion trampasense is replaced by Neohipparion eurystyle, Aphelops malacorhinus is replaced by Aphelops mutilus, Amebelodon floridanus is replaced by Amebelodon britti, Thinobadistes segnis is replaced by Thinobadistes wetzeli, and Pediomeryx hamiltoni is replaced by Pediomeryx hemphilliensis. Two new immigrants from the Old World first appear in the Hh2, the large bear Indarctos oregonensis and the otter Enhydritherium terraenovae (Tedford et al., 2004).
The Hh4 interval in Florida is very different from the Hh2. This is caused in part by the extinction event at the end of the Hh2 discussed above, in part by new immigrants from the Old World (Agriotherium, Eucyon, Lynx, PlesioguloCervidae, Miopetaurista), and in part by evolution of new genera and species from North American ancestors. Compared to western faunas, the Hh4 vertebrates of Florida actually show greater continuity with those of the early Hemphillian (Webb et al. 1995; 2008). For example, western Hh4 faunas lack Gomphotherium, Cormohipparion, protoceratids, and gelocids, while Pseudhipparion is found in the Hh4 of Oklahoma and Kansas, but not further west nor in Mexico. Western Hh4 are numerically dominated by one-toed species of horses such as Astrohippus stockii and “Dinohippus” mexicanus and three-toed species are rare, while in Florida Hh4 one-toed horses are far outnumbered by three-toed hipparions such as Nannippus aztecus, Cormohipparion emsliei, Neohipparion eurystyle, and Pseudhipparion simpsoni (Webb et al., 2008). Relatively brachydont herbivores, such as Tapirus, Pleiolama, Eocoileus, and Gomphotherium, were common in Florida during the Hh4, but very rare in the west.
Sources
- Text Author: Richard C. Hulbert Jr.
- Image Gallery: Natali Valdes
- Original Publication Date: June 18, 2015
- Last Edited On: August 2, 2016
References
Dalquest, W. W. 1983. Mammals of the Coffee Ranch Local Fauna, Hemphillian of Texas. The University of Texas at Austin, Texas Memorial Museum, Pearce-Sellards Series No. 38, 41 p.
Farlow, J. O., et al. 2001. The Pipe Creek Sinkhole Biota, a diverse late Tertiary continental fossil assemblage from Grant County, Indiana. American Midland Naturalist 145(2):367-378.
J. O. Farlow, R. L. Richards, R. C. Garniewicz, M. Greenan, W. R. Wepler, A. J. Shunk, G. A. Ludvigson, N. R. Shaffer, and A. L. Wenning. 2010. Occurrence and features of fossiliferous sediments of the Pipe Creek Sinkhole (late Neogene, Grant County, Indiana). Indiana Geological Survey Special Report 69:1-60. http://www.kgs.ku.edu/General/Personnel/klm/GAL/Farlow_etal_2010_PCS_monograph.pdf
Gustafson, E. P. 2012. New records of rhinoceroses from the Ringold Formation of central Washington and the Hemphillian-Blancan boundary. Journal of Vertebrate Paleontology 32(3):727-731.
Eshelman, R. E., and F. C. Whitmore Jr. 2008. Early Pliocene (late Hemphillian) land mammals from the Lee Creek Mine, Aurora, North Carolina. Pp. 17–38 in C. E. Ray, D. J. Bohaska, I. A. Koretsky, L. W. Ward, and L. G. Barnes (eds.), Geology and Paleontology of the Lee Creek Mine, North Carolina, IV. Virginia Museum of Natural History Special Publication 14, Martinsville.
Hulbert Jr., R. C. (ed.) 2001. The Fossil Vertebrates of Florida. University Press of Florida: Gainesville.
Hulbert Jr., R. C. and F. C. Whitmore, Jr. 2006. Late Miocene mammals from the Mauvilla Local Fauna, Alabama. Bulletin of the Florida Museum of Natural History 46(1):1-28.
Hulbert, R. C., S. C. Wallace, W. E. Klippel, and P. W. Parmalee. 2009. Cranial morphology and systematics of an extraordinary sample of the late Neogene dwarf tapir, Tapirus polkensis (Olsen). Journal of Paleontology 83(2):238-262.
Matthew, W. D., and R. A. Stirton. 1930. Equidae from the Pliocene of Texas. Bulletin of the Department of Geological Sciences, University of California Publications 19(17):349-396.
Reed, L. C, and O. M. Longnecker. 1932, The geology of Hemphill County, Texas: Austin, University of Texas Bulletin No. 3231, 98 p.
Schultz, G. E. 1990. Stop 16: late Hemphillian faunas of the Texas and Oklahoma Panhandles. Pp. 104-111 in T. C. Gustavson (ed.), Tertiary And Quaternary Stratigraphy And Vertebrate Paleontology Of Parts Of Northwestern Texas And Eastern New Mexico. Bureau of Economic Geology, University of Texas at Austin, Guidebook 24.
Tedford, R. H., L. B. Albright III, A. D. Barnosky, I. Ferrusquia-Villafranca, R. M. Hunt Jr., J. E. Storer, C. C. Swisher III, M. R. Voorhies, S. D. Webb, and D. P. Whistler. 2004. Mammalian biochronology of the Arikareean through Hemphillian interval (late Oligocene through earliest Pliocene epochs). Pp. 169-231 in M. O. Woodburne, (ed.), Late Cretaceous and Cenozoic Mammals of North America, Biostratigraphy and Biochronology. Columbia University Press, New York.
Webb, S. D. 2000. Evolutionary history of New World Cervidae. Pp. 38-64 in E. Vrba and G. Schaller (eds.), Antelopes, Deer, and Relatives: Fossil Record Behavioral Ecology, Systematics and Conservation. Yale University Press, New Haven.
Webb, S. D., R. C. Hulbert Jr., and W. D. Lambert. 1995. Climatic implications of large-herbivore distributions in the Miocene of North America. Pp. 91–108 in E. S. Vrba, G. H. Denton, T. C. Partridge, and L. H. Burckle (eds.), Paleoclimate and Evolution, with Emphasis on Human Origins. Yale University Press, New Haven.
Webb, S. D., R. C. Hulbert Jr., G. S. Morgan, and H. F. Evans. 2008. Terrestrial mammals of the Palmetto Fauna (early Pliocene, latest Hemphillian) from the Central Florida Phosphate District. Pp. 293-312 in X. Wang and L. G. Barnes (eds.), Geology and Vertebrate Paleontology of Western and Southern North America. Natural History Museum of Los Angeles County Science Series, Number 41.
Wood, H. E., et al. 1941. Nomenclature and correlation of the North American continental Tertiary. Bulletin of the Geological Society of America 52(1):1-48.