Quick Facts
Common Name: none
Enhydritherium terraenovae is a large, extinct otter capable of living in both freshwater and coastal marine habitats.
A nearly complete skeleton is on permanent public display at the Florida Museum of Natural History. It was collected at the Moss Acres Racetrack Site in Marion County.
Its presence in both Florida and California suggest it dispersed westward from the Gulf of Mexico into the eastern Pacific through the still open Panamanian Seaway. This predicts that its fossils should be recovered in late Miocene marine deposits in Panama.
Age Range
- Late Miocene to very early Pliocene epochs; early to late Hemphilian (Hh2-Hh4) land mammal age
- About 6.5 to 4.5 million years ago
Scientific Name and Classification
Enhydritherium terraenovae Berta and Morgan, 1985
Source of Species Name: The species name was formed by the combination of the Latin words terra meaning ‘earth’ or ‘land’ and novus meaning ‘new’, in reference to the species distribution in the New World, in contrast to closely related fossil otters known from Asia, Europe, and Africa.
Classification: Mammalia, Eutheria, Laurasiatheria, Carnivora, Caniformia, Arctoidea, Musteloidea, Mustelidae, Lutrinae
Alternate Scientific Names: none
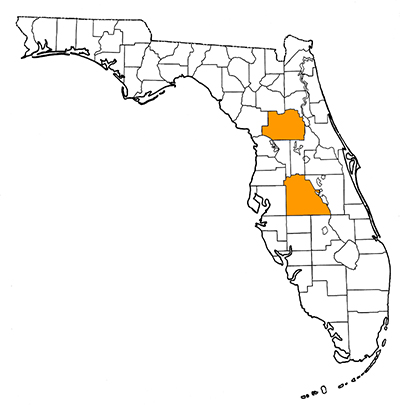
Overall Geographic Range
In addition to records from Florida, the species is also known from several locations in coastal southern California (Berta and Morgan, 1985). The type locality is the Palmetto Mine, Polk County, Florida.
Florida Fossil Occurrences
Florida fossil sites with Enhydritherium terraenovae:
- Marion County-Moss Acres Racetrack Site; Withlacoochee River 4A
- Polk County-Brewster Mine; District Grade Mine; Fort Green Mine; Fort Meade Mine (Gardinier); Whidden Creek Site in Fort Meade Mine (Gardinier); Palmetto Mine; Phosphoria Mine; Rockland Mine
Discussion
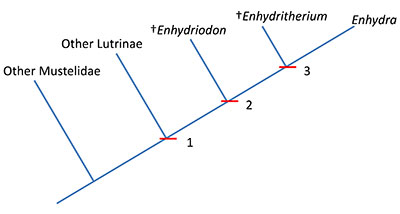
Otters, or members of the mustelid subfamily Lutrinae, fall into into two different groups based on the anatomy of their carnassial teeth, the upper fourth premolar (P4) and the lower first molar (m1). One group is called the “fish-eating” otters (Repenning, 1976; Lambert, 1997) and includes the extinct otter genus Satherium and the extant otter genera Lutra, Lontra, Pteronura, Amblonyx, and Aonyx. These otters are characterized by more blade-like carnassials, unlike the other group of otters, the bunodont otters. Bunodont otters possess non-blade-like carnassials with thick enamel and rounded cusps. The bunodont otters include the extant genus Enhydra (sea otters) as well as the extinct genera Enhydritherium and Enhydriodon (Lambert, 1997).
The genus Enhydritherium is thought to be closely related to the Enhydra and Enhydriodon, two genera found in late Miocene to early Pleistocene fossil sites of Europe, Asia, and Africa (Berta and Morgan, 1985; see Figure 2). This clade may be differentiated from all other members of the subfamily Lutrinae by the lack of the first premolar, a shorter rostrum, and bunodont cusps on the lower first molar (node 2, Figure 2). Enhydriodon is overall larger in size and is characterized by an “isolated” hypocone on the upper fourth premolar, which is different from Enhydritherium and Enhydra, which share several features including similar body size and the shapes of the upper fourth premolars and lower first molars (node 3, Figure 2). Enhydritherium may be distinguished from both Enhydriodon and Enhydra by the anterior and medial position of the protocone in the upper fourth premolar (Figure 3) and by the absence of the protostylid and presence of the metastylid on the lower first molar (Figure 4; Berta and Morgan, 1985; Lambert, 1997). Currently, Enhydritherium terraenovae is the only species in the genus.
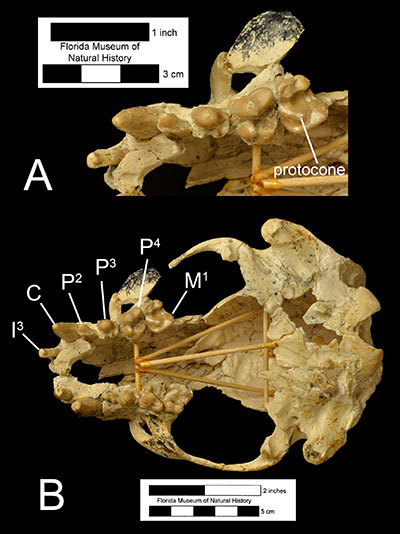
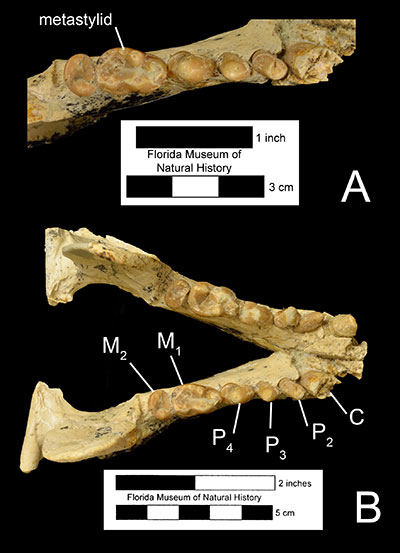
Enhydritherium terraenovae was at first primarily described from partial mandibles (Figure 5B-C; Berta and Morgan, 1985), although some isolated limb bones were tentatively referred to the species. A nearly complete skeleton of Enhydritherium terraenovae (UF 100000) was later found in 1987. It includes the skull, lower jaw, vertebrae, forelimbs, and hind limbs (Lambert, 1997). Before the discovery of UF 100000, Enhydritherium terraenovae had primarily been recovered from marine and nearshore fossil deposits in both Florida and California, leading Berta an Morgan (1985) to conclude that its ecology was similar to that of the modern sea otter, Enhydra. However, after the discovery of UF 100000 at the Moss Acres Racetrack site, which is believed to be a freshwater lake deposit in the middle of a grassland habitat, it has been hypothesized that E. terranovae was probably more terrestrial than previously believed, capable of moving inland to exploit freshwater habitats. Evidence from the postcranial fossils of E. terraenovae supports this hypothesis. Its limb proportions are more like those of the modern river otter Lontra and other “fish-eating” otters, which are more capable of terrestrial locomotion than the sea otter Enhydra. Furthermore, there is little evidence of swimming specializations in the hindlimb of E. terraenovae compared to Enhydra, which is more adapted for propulsive swimming using its hindlegs. In fact, E. terraenovae has a more robust greater tubercle on the humerus in comparison to Enhydra, which suggests that E. terraenovae primarily swam using its forelimbs. Although metapodial proportions suggest that E. terraenovae was probably less capable of terrestrial locomotion than Lontra, the lack of swimming specializations in the hindlimbs suggest they were still more terrestrially capable than Enhydra.
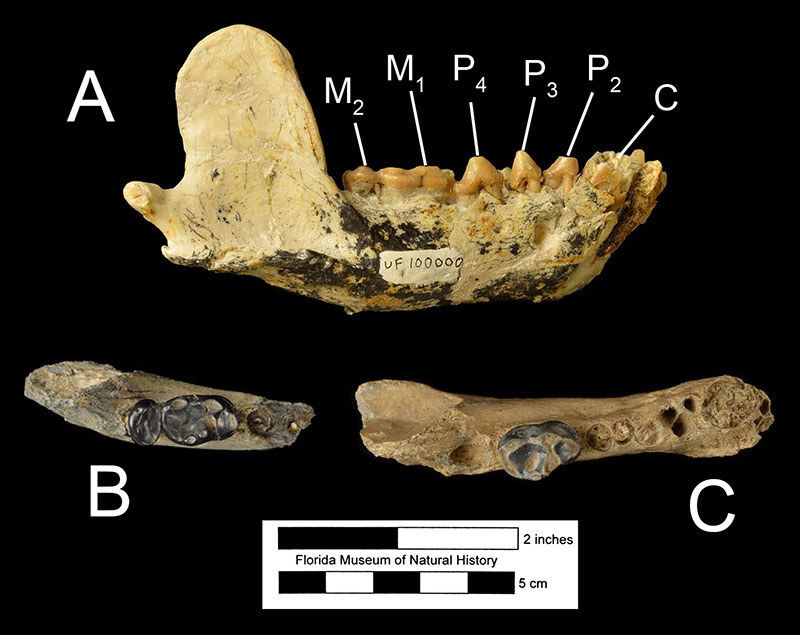
While the bunodont dentition of Enhydritherium terraenovae suggests that this species was adapted to eat tough foods (such crabs and shellfish) like Enhydra, the only extant otter with this type of dentition, Lambert (1997) hypothesizes that the diet of Enhydritherium terraenovae was not strictly limited to this food type following several lines of reasoning. Firstly, modern populations of Enhydra are not exclusively durophagous (tough-food eating), and some populations have been known to eat significant amounts of fish and cephalopods. Secondly, UF 100000 was found together with a large concentration of fish bones. This led Lambert (1997) to conclude that these fish bones were the stomach contents of UF 100000, as fish remains were otherwise rare at the site. Furthermore, molluscs thought to be common in freshwater habitats in the Miocene of Florida were probably too small (less than 3 cm or 1.2 inches in diameter) to exclusively sustain an animal the size of Enhydritherium terraenovae, which is hypothesized to have weighed between 22 and 45 kg (49–100 lbs).
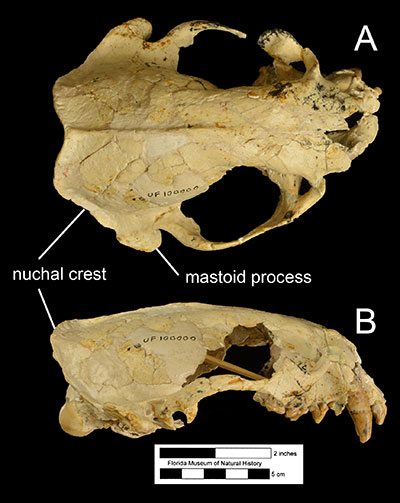
Lambert (1997) also suggested that Enhydritherium terraenovae was more reliant on fish for food than Enhydra, based on anatomical features. Firstly, its metacarpal proportions were more similar to those of modern fish-eating otters and unlike those of Enhydra. The hands of the bunodont otter Enhydra are specialized for procuring and handling food items, whereas modern fish-eating otters primarily procure food with their jaws. The specialization of the hands for food manipulation is also hypothesized to be the reason why Enhydra relies primarily on the hindlimbs for swimming. Secondly, the cranium of Enhydritherium terraenovae has a more well-defined nuchal crest and a long, robust mastoid process relative to modern otters. Because the nuchal crest and the mastoid process are the attachment sites of neck muscles, the robustness of these cranial features suggest relatively powerful neck muscles. The combined evidence of a lack of hindlimb swimming specialization, hand proportions more similar to modern fish-eating otters, and osteological evidence for powerful neck muscles, suggest that Enhydritherium terraenovae was well-adapted for catching fish and other moving prey with its jaws (Lambert, 1997).
Sources
- Original Author(s): Richard C. Hulbert, Jr. and Arianna Harrington
- Original Completion Date: February 13, 2013
- Editor(s) Name(s): Richard C. Hulbert, Jr., Natali Valdes, and Arianna R. Harrington
- Last Updated On: March 10, 2015
This material is based upon work supported by the National Science Foundation under Grant Number CSBR 1203222, Jonathan Bloch, Principal Investigator. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.
Copyright © Florida Museum of Natural History, University of Florida