by W. Mark Whitten, Florida Museum of Natural History, Gainesville FL and Colette C. Jacono, United States Geological Survey, Gainesville FL
Abstract
Marsilea ferns (water clover) are potentially invasive aquatic and wetland plants that are difficult to identify to species because of subtle diagnostic characters, sterile specimens, and unresolved taxonomic problems. We utilized DNA sequencing of several plastid regions to attempt to “fingerprint” Marsilea specimens from the southeastern U.S. to provide more accurate identifications. Currently, seven species are recognized from the eastern U.S., 6 of which have been found in Florida. Our data show that Florida specimens previously identified as M. oligospora (Jacono and Johnson 2006) are not true M. oligospora (native to the western U.S.), and instead may represent either an unknown introduced species or an undescribed native species.
Molecular data conflict with the morphological characters for distinguishing M. vestita, M. macropoda, and M. nashii. The molecular data reveal two clades within this species complex: 1. A western U.S/Mexican clade; and 2. an eastern U.S./coastal plain clade. This DNA/morphology discordance suggests that these taxa may have hybridized extensively or may represent only one variable species; either case warrants reexamination of the species level-taxonomy of the group. DNA data clearly distinguish all other species of introduced Marsilea, and DNA sequencing is valuable as a tool for identification of sterile Marsilea specimens.
Introduction
Marsilea (Water-Clover) is a genus of aquatic ferns that is increasingly being found as invasive in Florida and the Southeast. Marsilea (ca. 50 spp.) occur worldwide as two ecological types 1) true aquatic species with glabrous leaves that inhabit permanent water bodies and 2) semi-aquatic species with hairy leaves that prefer fluctuating wetland habitats and prevail through seasonal extremes in wet and dry periods. Florida’s lakes and wetlands provide habitat for both types and currently host at least six species of both ecological types, all of which have been hypothesized as introduced.
The Water-Clovers bear few dependable morphological characters on which to base traditional identification. Morphological plasticity is widespread, and sporocarps, an important identification feature, are commonly absent in field populations. Because identification of Marsileabased upon morphology is so problematic, aquatic plant managers need more reliable tools for identification. Without objective methods of identification, managers cannot make decisions about the probable identity and origin of introduced species and how to manage populations. In this study, we use DNA sequences of four plastid regions (rbcL, rps4, the rps4-trnS spacer, and the trnL-F spacer) to aid in identification of Marsilea specimens. The recent molecular phylogeny of Marsilea (Nagalingum et al., 2007) provides a phylogenetic framework for comparison of sequences in GenBank with those from additional specimens.
Several populations of Marsilea in Florida (far from natural, western U.S. distributions) have uncertain identities. Of primary interest are historical specimens, collected in the early 1890s, the identification of which has vacillated from M. vestita, an introduction from the western U.S. (Ward and Hall 1976) to M. ancylopoda, a rare and potentially extinct native species (FNA 1993). Plants in recently reported populations in three central Florida counties appearing identical to plants from the historic sites were identified as M. oligospora, a semi-aquatic North American species endemic to the northern fringe of the Great Basin (Jacono and Johnson 2006). Nevertheless, variation was noted between the Florida and the Great Basin material and it was difficult for the authors to speculate how a geographically restricted plant with no known economic value may have become established in central Florida over 100 years ago.
At the same time, invasive plant managers are interested in initiating herbicide practices on pest populations of Marsilea. One site of particular concern is Emeralda Marsh, a 1,500 acre restoration unit bordering Lake Griffin, Lake County, FL where M. oligospora appears to be increasing in wetlands where the naturally fluctuating hydrologic regime is being restored. Molecular data may provide a more confident determination of these specimens and help to resolve whether these populations are introduced weeds to be targeted for herbicide treatment or if they are a native component of natural wetlands.
Our specific objective is to provide molecular evidence to support or refute morphological identification and region of origin of the central Florida populations. Such evidence will provide a more sound basis for future pest plant listing and for responsible management practices. However, the laboratory procedures to be developed are a modest investment with wider application than identification of a single species. The method will serve as a screening protocol for well established, new and potentially invasive species of this difficult and typically sterile genus. In this project we surveyed all known populations of Marsilea within Florida using several molecular markers and compared them to all U.S. and Caribbean species, as well as Marsilea species common in the aquatic plant trade that are becoming established in the Southeast. These data will provide a baseline for evaluating the identity and nativity of species in Florida and for distinguishing future introductions of Marsilea.
Methods
Appendix 1 presents a list of voucher specimens sampled. Samples were taken from herbarium specimens (with permission) from various herbaria. Because Florida collections of M. oligospora were hypothesized to be introductions from the western US (Jacono and Johnson 2006), we included specimens from several western herbaria (from the vicinity of the type locality). Leaf samples (ca. 25 mm2) were ground using a tissue mill and extracted using a modified version of the 2x CTAB procedure of Doyle and Doyle (1987) with exclusion of beta-mercaptoethanol and inclusion of 5 units of proteinase K to improve yield and quality of DNA. Primers for rbcL were designed to allow amplification and sequencing in two overlapping pieces, facilitating amplification from degraded total DNAs. Primers for rbcL are: rbcL F ATGTCACCACAAACAGAGACTAAAGC; rbcL intF TGAGAACGTAAACTCCCAACCATTCA; rbcL intR CTGTCTATCGATAACAGCATGCAT; and rbcL R GCAGCAGCTAGTTCCGGGCTCCA. The rps4 exon and the adjacent rps4-trnS spacer were amplified in one piece using the primers rps4F ATGTCCCGTTATCGAGGACCT and rps4R TACCGAGGGTTCGAATC. Primers for the trnL-F spacer (primers E&F) were those of Nagalingum et al. (2007). All amplifications utilized Sigma Jumpstart Taq polymerase and reagents (Sigma-Aldrich, Inc.) in 25 μl reactions with 3.0 mM MgCl2. Thermocycler conditions were: 94 C for 3 minutes followed by 37 cycles of 94 C for 30 s, 56 C for 30 s, 72 C for 2 min, with a final extension of 3 min at 72 C. PCR products were sequenced in both directions, using the Big Dye Terminator reagents on an 3130 automated sequencer following manufacturer’s protocols (Applied Biosystems, Inc.). Electropherograms were edited and assembled using Sequencher 4.8 (Genecodes Inc., Ann Arbor, Michigan, USA), and the resulting sequences were aligned manually. All sequences will be deposited in GenBank prior to publication. Analyses were performed using PAUP* version 4.0 b10 (Swofford, 2003) with Fitch parsimony (equal weights, unordered characters, ACCTRAN optimization and gaps treated as missing data. Heuristic searches consisted of 1000 random taxon addition replicates of subtree-pruning-regrafting (SPR) and keeping multiple trees (MULTREES) with the number of trees limited to 10 per replicate to minimize extensive swapping on islands with many suboptimal trees.
Results and Discussion
Figures 1-3 presents a randomly chosen single most parsimonious tree. The tree combines data in GenBank from the world wide taxonomic sampling of Nagalingum et al. (2007) with data generated from this study (Appendix 1). Individuals noted in full Latin binomial are from GenBank data. Individuals noted by “M.” followed by a species name and three digit number and locality, usually a state and county, are primarily U.S. collections sampled by us in this study.
The Florida collections of Marsilea fall into five distinct clades within the tree; these clades are highlighted in gray and labeled Clades A-E.
The DNA data revealed that numerous specimens sampled in this study (especially sterile ones) were misdetermined. DNA analysis was especially effective in clarifying the identification of sterile specimens of both North American and introduced origin and was equally effective with aquatic and semiaquatic ecotypes.
Little or no genetic variation among geographically distant introductions of the same species (eg. M. minuta, M. hirsuta) suggests a wide reaching water garden industry as the source of North American material.
M. mutica (clade A; Figure 1) is clearly distinct from all other taxa in terms of DNA distance. This parallels its morphology which is unique from all other taxa for its two-toned leaflets and petioles inflated at the apex to function as air bladders for floating leaves. Indigenous to Australia and New Caledonia, M. mutica may be the most popular species in the water garden trade. The SE US specimens plus one from Oklahoma form a clade distinct from specimens from Arizona and Virginia. This is suggestive of at least two distinct geographic origins for material introduced into the US.
Clade B (Figure 2) includes M. hirsuta, an Australian species introduced into Florida. Three Australian hairy leaved species (M. drummondii, M. angustifolia and M. hirsuta) form a clade distinct from the Australian glabrous leaved species, M. mutica. DNA data fail to distinguish M. hirsutafrom M. angustifolia. Morphologically, M. angustifolia differs from M. hirsuta in having smaller and more elongated leaves and smaller sporocarps (Aston 1973). These characters, however, are typically considered insufficient for species distinction within the genus Marsilea (Launert 1968). Our DNA data suggests that the taxonomic boundaries of these two species should be reexamined.
Clade C (Figure 2) consists of M. minuta (plus M. crenata, M. fadeniana). Samples from introductions in the southeastern U.S. appear more closely related to M. minuta from India, and to M. crenata, from Thailand and Indonesia than to M. minuta from Africa. A population of M. minuta introduced to Trinidad since at least 1982 is discrete from the southeast material and more closely related to M. minuta and M. fadeniana from Africa. As presently circumscribed, M. minuta is a geographically widespread species that needs further taxonomic clarification. Although not tested in this study, M. crenata, which has been reported as introduced to Hawaii is probably the same entity as M. minuta in the SE US. Johnson (1986) suggested that M. quadrifolia, long established in the northeastern U.S., may be close to M. minuta, but, based on the one specimen from Japan sampled in this study, this would not be the case. M. quadrifolia is the most temperate of the Marsilea species. Generally, introductions of M. quadrifolia are from earlier dates and are believed to have originated from Europe. Paradoxically, in Europe it is now a Red Data species, listed for its rarity and vulnerability to extinction.
Clade D (Figure 2) includes Florida specimens previously determined as M. oligospora by Jacono (thought to represent an introduction from the western US), but DNA data show that these specimens do not match any material tested or in GenBank. They are clearly distinct from specimens of “true” M. oligospora (Figure 3) from the western US (close to the type locality). The DNA data demonstrates that it is a distinct species, yet related to M. ancylopoda, a widely distributed neotropical species with which it was earlier misidentified (FNA 1993). The geographic range of this M. aff. oligospora plus the existence of Florida collections from the 1890s suggests an alternative hypothesis: that it may be a native species. Although the very recent expansion of the species at artificial and disturbed habitats seems to dispute this assumption, only greater sampling of species in the Caribbean and tropical Americas might reveal the true identity of M. aff. oligospora in Florida.
Clade E (Figure 3) consists exclusively of plants from the U.S. Gulf Coast and Florida, and includes specimens determined as either M. vestita, M. macropoda, or M. nashii (from GenBank). Although well accepted as distinct North American species, the morphological distinctions separating vestita and macropoda clearly do not agree with the molecular data presented here. The correct name for plants in this clade (probably M. macropoda or M. nashii) is yet to be determined, and will require comparison of sequence data with specimens from the type localities of these two species. Comparison with specimens noted earlier as putative hybrids between M. vestita and macropoda in Texas (Johnson 1986) should also prove informative. The demarcation of specimens into Clade E introduces the scenario that plants previously labeled vestita and macropoda and considered western NA introductions into Florida, may in fact represent an independent entity (or hybrid) that could be native in Florida. If this were the case, it may serve as indirect support for the Clade D entity (M. aff. oligospora) occurring as native to Florida.
Clade F (Figure 3) consists of specimens of M. vestita and M. oligospora from the western US and Mexico, and suggests that species boundaries and taxonomy of these taxa need to be reevaluated. The Hawaiian species M. villosa and a specimen determined as M. fourneiri also fall in Clade F. Clearly, the taxonomic relationships of M. vestita, its subspecies, and related species need to be reexamined. One possible explanation for the geographic distinction between clades E and F is that these species have hybridized (resulting in a single chloroplast type because of uniparental inheritance of chloroplasts); this hypothesis could be evaluated by sequencing of a suitable nuclear marker that is biparentally inherited. However, we know of no experimental or molecular evidence that Marsileas do hybridize in nature.
Conclusions/Recommendations
In light of the difficulties inherent to morphological identification of Marsilea, resource managers can opt for DNA analysis as a reliable tool in species identification. DNA analysis effectively determined sterile specimens of North American and introduced species at a minimal cost per sample. Results from DNA analysis also revealed a low level of genetic diversity among collections of geographically distant introductions of the same species, which indicates a wide reaching industry as a source of the eastern hemisphere species M. minuta, M. hirsuta, and M. mutica.
DNA results further indicate that Florida M. aff. oligospora should not be regarded as an introduced species nor targeted for eradication until more is known about its identity. If the entity represents an undescribed native species, its increase at restoration marshes could be a sign of the return of more natural hydroperiods and ecological dynamics.
Discrepancies demonstrated by DNA analysis in the taxonomic relationships among specimens labeled M. vestita or M. macropoda has in the past been addressed by designation of morphological subspecies. In light of the geographical patterns found in our study, reexamination of this species group may reveal that a hybrid of the two, or a third species, could be native in Florida. If this were the case, it may further support the nativity of M. aff. oligospora in that state.
Literature Cited
- Aston, Helen 1973. Aquatic Plants of Australia: A guide to the identification of the aquatic ferns and flowering plants of Australia, both native and naturalized. National Herbarium of Victoria, Melbourne University Press, Melbourne.
- Chase, M.W., R. S. Cowan, P. M. Hollingsworth, C. van den Berg, S. Madriñán, G. Petersen, O. Seberg, T. Jørgsensen, K. M. Cameron, M. Carine, N. Pedersen, T. Hedderson, F. Conrad, G. Salazar, J. Richardson, M. Hollingsworth, T. Barraclough, L. Kelly and M.Wilkinson. 2007. A proposal for a standardised protocol to barcode all land plants. Taxon 56(2):295-300.
- Doyle, J. J. and J. L. Doyle. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin 19: 11-15.
- (FNA) Flora of North America Editorial Committee (eds.). 1993. Flora of North America North of Mexico. Volume 2. Pteridophytes and Gymnosperms. Oxford University Press, New York.
- Jacono, C. C. and D. M. Johnson. 2006. Water-clover Ferns, Marsilea, in the Southeastern United States. Castanea 71(1):1-14.
- Johnson, D.M. 1986. Systematics of the new world species of Marsilea (Marsileaceae). Systematic Botany Monographs 11:1-87.
- Launert. E. 1968. A monographic survey of the genus Marsilea Linnaeus. Senckenbergiana Biologica 49:273-315.
- Nagalingum, N. S., H. Schneider, and K. M. Pryer. 2007. Molecular phylogenetic relationships and morphological evolution in the heterosporus fern genus Marsilea. Systematic Botany 32:16-25.
Figure 1. Base of single most parsimonious tree for combined molecular data set.
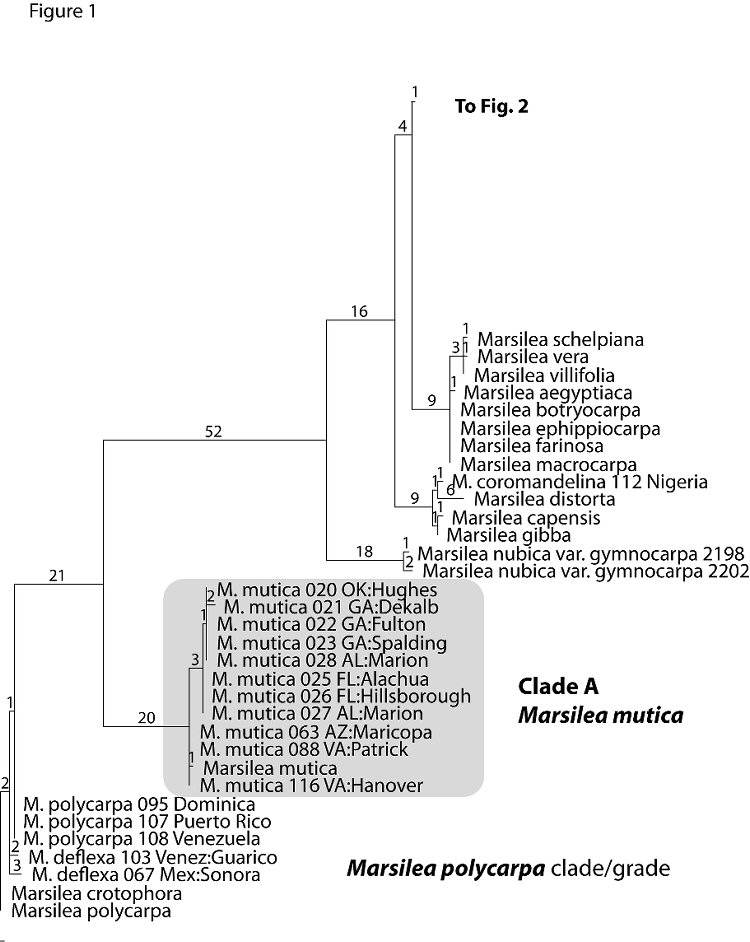
Figure 2. Mid-portion of single most parsimonious tree for combined molecular data set.
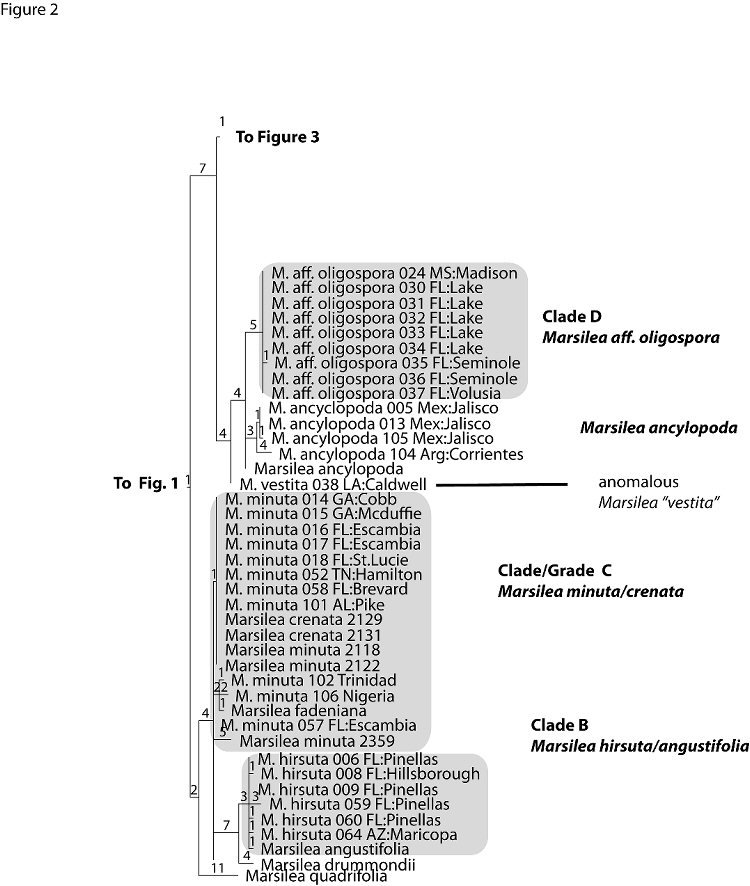
Figure 3. Terminal clades of single most parsimonious tree for combined molecular data set

Appendix 1. Specimens sampled for DNA analysis in this study.
DNA numbers (column 1) correspond to three-digit numbers following taxon names in figures 1-3. Some specimens failed to amplify or sequence cleanly, and are not represented in the tree.
| DNA number | Taxon | Collector | Herbarium | Herb acc . | Locality | Locality detail | mo-day-yr |
|---|---|---|---|---|---|---|---|
| 5 | Marsilea ancylopoda | Lott 3321 | FLAS | 184721 | Mexico: Jalisco | "Mpio. La Huerta: Las Alamandas, ca. 26 km NW of Estacion de Biologia Chamela on the Puerto Vallarta-Barra de Navidad (Mex. 200) hwy, and 3 km W of the village of Quemaro." | 1/16/91 |
| 6 | Marsilea hirsuta | Beckner 2619 | FLAS | P8932 | FL: Pinellas | "St. Petersburg, 46th St. and Fairfield Ave. S." | 8/13/83 |
| 7 | Marsilea macropoda | Ward 7623 | FLAS | P7699 | TX: San Patricio | "9 mi NE of Sinton, near west end of Welder Wildlife Refuge." | 5/1/71 |
| 8 | Marsilea hirsuta | Jacono 257 | FLAS | 219917 | FL: Hillsborough | "Northwest Mitigation Bank, Area C, off S. Mobley Rd, ca. 3 mi. SW of Citrus Park." | 4/22/01 |
| 9 | Marsilea hirsuta | Jacono 235 | FLAS | 219921 | FL : Pinellas | "St. Petersburg, 46th St. and Fairfield Ave. S." | 3/31/01 |
| 10 | Marsilea macropoda | Burkhalter 8846 | FLAS | P8925 | AL: Mobile | Mobile; SE corner of Conception St. and Texas St. | 6/11/83 |
| 11 | Marsilea macropoda | Burkhalter 5672 | FLAS | P8409 | AL: Mobile | Mobile; SE corner of Church Street and Washington Ave. | 12/20/77 |
| 12 | Marsilea macropoda | McAlpin s.n. | FLAS | P9864 | FL: Sarasota | "Banks of drainage ditch along 51 Street, just east of Rt. 301." | 6/15/87 |
| 13 | Marsilea ancylopoda | E. Lott 3321 dupe of 005 | FLAS | 184721 | Mexico: Jalisco | "Mpio. La Huerta: Las Alamandas, ca. 26 km NW of Estacion de Biologia Chamela on the Puerto Vallarta-Barra de Navidad (Mex. 200) hwy, and 3 km W of the village of Quemaro." | 1/16/91 |
| 14 | Marsilea quadrifolia | J. Allison 8001 | FLAS | 180750 | GA: Cobb Co. | "Ca. 0.7 mil WNW of Austell, W of Thornton Rd. and N of Garrett Rd.; Powder Springs Creek, near mouth confluence with Sweetwater Creek." | 9/18/93 |
| 15 | Marsilea minuta | J. Germann s.n. | FLAS | 219925 | GA: McDuffie Co. | "Thompson, pond near." | 6/1/00 |
| 16 | Marsilea minuta | Jacono 394 | FLAS | 219922 | FL: Escambia | "SW of Pensacola, N of Bayou Grande, Grundy St. and Lowndes Ave., NW side of intersection." | 2/28/02 |
| 17 | Marsilea minuta | Burkhalter 13304 | FLAS | 179393 | FL: Escambia | Near Pensacola; NW corner of Lownde Ave. and Grundy St. | 6/19/92 |
| 18 | Marsilea minuta | Jacono 498 | FLAS | 219924 | FL: St. Lucie | "Port St. Lucie, cultivated from material in canal." | 3/13/02 |
| 19 | Marsilea minuta | Jacono 512 | FLAS | 219923 | FL: Sarasota | Cultivated material originally from Marie Selby Botanical Garden. | 11/1/02 |
| 20 | Marsilea mutica | Jacono 503 | FLAS | 219936 | OK: Hughes | "Cultivated from Holdenville, Stroup Park pond." | 10/1/01 |
| 21 | Marsilea mutica | Jacono 362 | FLAS | 219931 | GA: Dekalb | "Davidson-Arabia Mountain Nature Preserve, lake off Rockland Rd." | 11/5/01 |
| 22 | Marsilea mutica | Cooper s.n. | FLAS | 219932 | GA: Fulton | "Palmetto; farm pond off Hutchinson Ferry Rd., ca. 4 mi from Hwy 29." | 10/24/01 |
| 23 | Marsilea mutica | Jacono 361 | FLAS | 219933 | GA: Spalding | Shoal Creek subdivision Lake. | 11/5/01 |
| 24 | Marsilea mutica | Kirk s.n. | FLAS | 219934 | MS: Madison | "Lake Lorman, SW of Hwy 463." | 12/11/01 |
| 25 | Marsilea mutica | Davis 1208 | FLAS | 215625 | FL: Alachua | Gainesville; Kanapaha Botanical Gardens. | 4/2/05 |
| 26 | Marsilea mutica | Rodgers s.n. | FLAS | 207131 | FL: Hillsborough | "Tampa: Lake Magdalene, N of Fletcher Ave., E of Dale Mabry." | 7/2/02 |
| 27 | Marsilea mutica | Greene s.n. | FLAS | 219928 | AL: Marion | "Hamilton, farm pond, S of John Craft Hwy, N of CR90." | 8/13/01 |
| 28 | Marsilea mutica | Jacono 502 | FLAS | 219930 | AL: Marion | "Cultivated from Hamilton farm pond, S of John Craft Hwq, N of CR 90." | 11/19/02 |
| 29 | Marsilea oligospora | Anderson 1299 | FLAS | P8835 | UT: Cache | "Dry Lake, 4 air miles NE of Brigham City." | 7/9/58 |
| 30 | Marsilea oligospora | Jacono 508 | FLAS | 219938 | FL: Lake | "Orange Bend, E side of road intersecting N of E Treasure Island Rd." | 12/8/02 |
| 31 | Marsilea oligospora | Jacono 510 | FLAS | 219940 | FL: Lake | "Emeralda Marsh, Treasure Island Entrance, St. Johns River Water Management Area." | 12/8/02 |
| 32 | Marsilea oligospora | Jacono 511 | FLAS | 219941 | FL: Lake | "Emeralda Marsh, W of Hwy 452; diked impoundment of Lake Griffin." | 12/8/02 |
| 33 | Marsilea oligospora | Jacono 571 | FLAS | 219942 | FL: Lake | Orange Bend. E side of road intersecting N of E Treasure Island Rd. | 5/1/04 |
| 34 | Marsilea oligospora | Jacono 572 | FLAS | 219943 | FL: Lake | "Emeralda Marsh, W of Hwy 452 Treasure Island Entrance of St. Johns Water Management District lands." | 5/1/04 |
| 35 | Marsilea oligospora | Brinson s.n. | FLAS | P9670 | FL: Seminole | "Roadsid ditch to Soldier's Creek, 2.5 miles west of Lake Jessup, west of SR 17/92" | 4/16/86 |
| 36 | Marsilea oligospora | Jacono 177 | FLAS | 219944 | FL: Seminole | "Soldier Creek, W side Hwy 17/92, approx. 6 air miles S of Sanford and 1.75 air miles W of Lake Jessup, just S of sign to ""Big Tree Park""." | 10/28/00 |
| 37 | Marsilea oligospora | Meisenburg s.n. | FLAS | 219414 | FL: Volusia | "Ormond Beach, 3 Wild Cat Lane, retention pond mitigation area." | 7/5/06 |
| 38 | Marsilea vestita | Raymond & Painter 81 | FLAS | P8220 | LA: Caldwell | Immediately off US 165 at Riverton. | 7/16/74 |
| 39 | Marsilea vestita | "Thomas 114,754" | FLAS | 171536 | LA: Caldwell | West bank of Ouachita River at Columbia Recreation Area west of US 165 at Columbia. | 11/21/89 |
| 40 | Marsilea vestita | Brodie s.n. | FLAS | 219946 | FL: Collier | "Immokalee, 618 9th St. S, E side of ditch." | 1/15/03 |
| 41 | Marsilea vestita | Jacono 619 | FLAS | 219948 | FL: Collier | "Cutivated in Gainesville from Immokalee, 618 9th St. S, E side of ditch." | 5/12/03 |
| 42 | Marsilea vestita | Anderson 7356 | FLAS | P9104 | FL: Franklin | "Apalachicola, Ave. K near 9th Street." | 6/19/84 |
| 43 | Marsilea vestita | Anderson 7625 | FLAS | P9105 | FL: Franklin | "Apalachicola, West side of N. Market Street (just N of Avenue I)" | 9/20/84 |
| 44 | Marsilea vestita | Jacono 183 | FLAS | 219949 | FL: Franklin | "Apalachicola, W of N Market St., N of Ave. F, S of Dr. Martin Luther King Jr. Ave." | 11/18/00 |
| 45 | Marsilea vestita | Jacono 504 | FLAS | 219950 | FL: Franklin | "Cultivated from Apalachicola, W of N Market St., N of Ave. F, S of Dr. Martin Luther King Jr. Ave." | 4/18/01 |
| 46 | Marsilea vestita | Hall 414 | FLAS | P7953 | FL: Hillsborough | Originally brought from New Orleans; weed in flower bed at 1407 W. College Ave. in Ruskin. | 9/25/75 |
| 47 | Marsilea vestita | D. Thomas & E. Sundell 167474 | TENN | AR: Lincoln | Huff Island Park along the Arkansas River at Joe Hardin Lock and Dam at the edn of Ark. 111 north of Grady; sect. 34 | 9/15/00 | |
| 48 | Marsilea vestita | D. Thomas 114172 | TENN | LA: Caldwell | Oachita River east of Copenhagen and La. 849 and south of Columbia; along sandbars and riverbank. | 10/31/89 | |
| 49 | Marsilea vestita | D. Thomas & C. M. Allen | TENN | LA: St. James | Beside La.44 and the Miss. River Levee at Dravco Materials Sunshine Yard about 2.1 miles North of Convent | 5/22/90 | |
| 50 | Marsilea vestita | J. Taylor 29759 | TENN | OK: Johnston | Headwaters area on Tishomingo National Wildlife Refuge | 8/28/80 | |
| 51 | Marsilea macropoda | G. Landry & S. Holder 7832 | TENN | LA: Orleans | New Orleans; Abandoned lot at corner of Bienville and N. Salcedo Street | 3/15/81 | |
| 52 | Marsilea minuta | J.T. Beck 3867 | TENN | TN: Hamilton | "Chattanooga: Lake Junior, near intersection of Hwy 153 and Amnicola Hwy Pond margins" | 10/18/00 | |
| 53 | Marsilea macropoda | K. Bradley 1858 | USF | 233213 | FL: Collier | "Immokalee: At west end of 9th Street, west of US 29" | 7/4/98 |
| 54 | Marsilea macropoda | B. McAlpin sn | USF | 192158 | FL: Sarasota | "Drainage ditch along 51st Street, just east of Rt. 301" | 6/15/87 |
| 55 | Marsilea macropoda | B. McAlpin sn | USF | 233128 | FL: Collier | "Immokalee: At west end of 9th Street, west of US 29." | 4/9/99 |
| 56 | Marsilea vestita | A. Gholson et al. 11253 | USF | 198702 | FL: Franklin | "Appalachicola: West side Market Street, 0.2 mi north from junction with Avenue F" | 4/5/85 |
| 57 | Marsilea minuta | J. Burkhalter 13304 | USF | 206485 | FL: Escambia | Near Pensacola; NW corner of Lownde Ave. and Grundy St. | 6/19/92 |
| 58 | Marsilea minuta | Jacono 667 | USF | 242904 | FL: Brevard | South Central Wastewater Treatment Plant; cult. In Gainesville FL | 6/9/06 |
| 59 | Marsilea hirsuta | J. Beckner 2619 | USF | 171665 | FL: Pinellas | "St. Petersburg, 46th St. and Fairfield Ave. South" | 10/1/83 |
| 60 | Marsilea hirsuta | R. Wunderlin et al. 10365 | USF | 191296 | FL: Pinellas | St. Petersburg; Fairfield Ave. S and 46th Stree South | 3/3/87 |
| 61 | Marsilea sp | C. Clarkson sn | USF | 200757 | FL: Hardee | Approx. 3 mi south of Zolfo Springs | 7/3/89 |
| 62 | Marsilea vestita | no collector | USF | 201283 | FL: Highlands | "Seabring; Billy Bowlegs Creek, Highlands Hammock State Park" | 1/1/71 |
| 63 | Marsilea mutica | D. Damrel 2429 | ASU | 234980 | AZ: Maricopa | "Arizona State University, Desert Arboretum Park" | 8/2/99 |
| 64 | Marsilea hirsuta | L. Makings 2090 | ASU | 260430 | AZ: Maricopa | Scottsdale; backyard lily pond | 6/5/05 |
| 65 | Marsilea mollis | G. & M. Hatschbach 54145 | ASU | 203441 | Brazil: MG | "Rio Sao Francisco, mun. Buritizeiro" | 6/9/90 |
| 66 | Marsilea vestita | R. Felger 85-484 | ASU | 267754 | MX: Sonora | San Jose de Guaymas | 3/6/85 |
| 67 | Marsilea deflexa | Steinman et al. 93-373 | ASU | 194802 | MX: Sonora | "Laguna Barochipa on the coast of the Gulf of California, ca. 3.3 km sse of Cmahuiroa." | 11/23/93 |
| 68 | Marsilea vestita | J. Rebman & H. Lopez 2608 | ASU | 203225 | MX: Baja California | "Rancho Las Chichihuas; between Ensendada and Rosarito along Mex. Rt. 1, south of La Mision." | 4/15/94 |
| 69 | Marsilea vestita | S. Friedman & K. Johnson 456-94 | ASU | 206907 | MX: Sonora | "0.8 km n of Diez de Abril at Arroyo Muerto and Powerline road, 5.25 km n of Melchor Ocampo" | 10/20/94 |
| 70 | Marsilea mollis | G. Ferguson 118 | ASU | 116486 | MX: Chihuahua | "Laguna Arareco, 7.5 km by road SE Creel on rd. to Cusarare" | 8/29/81 |
| 71 | Marsilea mexicana | J. Hernandez sn | ASU | 215353 | MX: Zacatecas | 2 km n of Valparaiso | 12/24/84 |
| 72 | Marsilea fourneieri | J. Rebman 1684 | ASU | 207533 | MX: Baja California Sur | Rio Commundu; 64 km n of Ciudad Insurgentes and 13.7 miles east of Rancisco Villa | 4/18/93 |
| 73 | Marsilea vestita | E. Wise 1716 | ASU | 146549` | CA: San Luis Obisbo | Santa Margarita Lake | 6/30/83 |
| 74 | Marsilea vestita | R. Worthington 21894 | ASU | 257286 | NM: Luna | "West edge of West Potrillo Mts. At edge of mts., 12 mi E of Columbus in playa roadside ditch" | 4/17/93 |
| 75 | Marsilea vestita | D. Keil 18024 | ASU | 135889 | CA: San Luis Obisbo | "Just S of Creston, W of O'Donovan Road, on fringes of small reservoir." | 10/23/83 |
| 76 | Marsilea vestita | L. McGill 7203 | ASU | 262038 | AZ: Cochise | Willcox; flood drainage channel across N 3rd Ave. between Stewardt St. and Historic Willcox Cemetary. | 11/9/00 |
| 77 | Marsilea mollis | W. T. Johnson sn | ASU | 133244 | AZ: Coconino | "Baker Lake, Mogollon Rim, jct. state hwy 87 and Forest Road 218A" | 6/29/84 |
| 78 | Marsilea vestita | G. Marrs-Smith 1211 | ASU | 135685 | AZ: Cochise | "San Bernardino Ranch, 18 mi E of Douglas, north part of Black Draw." | 9/21/81 |
| 79 | Marsilea mollis | D. Damrel 627-A | ASU | 241765 | AZ: Gila | "Sierra Ancha Mts, off FSRD 682 and 486 intersection, at Indian Camp Tank." | 9/20/02 |
| 80 | Marsilea mollis | T. Wright & M. Baker 93-102 | ASU | 202936 | AZ: Yaupai | "NW of Prescott, 1.6 km NW of Pinetop Mtn, 1.2 km SE of Hunt Tank" | 6/18/93 |
| 81 | Marsilea mollis | J. Collins sn | ASU | 124092 | AZ: Cochise | "San Rafael, Rafael Valley" | 9/23/82 |
| 82 | Marsilea mollis | E. Lehto 24541 | ASU | 111936 | AZ: Coconino | "Foxborough Lake, Schnebly Mill road." | 6/26/80 |
| 83 | Marsilea mollis | M. Baker 8595 | ASU | 116549 | AZ: Coconino | "Twenty-Nine Mile Lake, ca. 5 miles due northo f Pine" | 8/16/91 |
| 84 | Marsilea mollis | L. McGill 6860 | ASU | 210327 | AZ:Cochise | "Turkey Creek Ranch, est drainage from Chiricuahua Mts into Sulphur Spring Valley; 1/4 mi N of Hwy 181 E at milemarker 49.5" | 10/1/96 |
| 85 | Marsilea mollis | M. Windham 0114D | ASU | 113526 | AZ: Coconino | "Coconino National Forest, sw shore of Foxboro Lake 3.29 km ENE of Schnebly Hill Vista and 1.72 km WSW of Mason Tank." | 10/20/79 |
| 86 | Marsilea mollis | Pinkava sn | ASU | 216827 | AZ:Coconino | Foxboro Lake | 5/4/96 |
| 87 | Marsilea vestita | R.D. Thomas 114754 | VPI | 82735 | LA: Caldwell | Along west bank of Oachita River at Columbia Recreation area west of US 165 at Columbia | 11/21/89 |
| 88 | Marsilea mutica | M. Robinette sn | VPI | 100854 | VA: Patrick | Persistent weed in pond in vicinity of Critz | 5/25/05 |
| 89 | Marsilea vestita | C.R. Bjork 6868 | ID | 122740 | ID: Nez Perce | "Clearwater River, 0.5 km east of the mouth of Catholic Creek." | 9/23/02 |
| 90 | Marsilea vestita | C.R. Bjork s.n. | ID | 122926 | ID: Nez Perce | Ca. 2 miles east of Spalding along Clearwater River; Shallow seasonal pools below Hwy 12. | 10/1/99 |
| 91 | Marsilea vestita | A. Tiehm 15267 | ID | 139382 | NV: Washoe | "Virginia Mts., NW side of Spanish Flat Reservoir at end of Dry Valley Creek." | 7/18/06 |
| 92 | Marsilea vestita | C.R. Bjork 2404 | ID | 134760 | OR: Malheur | Snake River Plains Bioregion; Interstate 84 crossing of the Snake River. | 8/1/96 |
| 93 | Marsilea oligospora | C.R. Bjork 3916 | ID | 124971 | ID: Owyhee | "Duck Valley Indian Reservation; about 11 mi. S of Riddle, creek along Hwy 51." | 8/1/98 |
| 94 | Marsilea oligospora | F.D. Johnson sn | ID | 80216 | ID: Idaho | "Swartz Bar, lower Salmon River about 10 mi. NW of Whitebird." | 8/9/80 |
| 95 | Marsilea polycarpa | S.R. Hall & L. Phillipe 28868 | MO | 5161408 | Dominica: St. Andrews | "East end of Melville Hall airport, cow pasture near dump area at coast" | 2/23/97 |
| 96 | Marsilea vestita | N. & P. Holmgren | UTC | 241370 | NV:Elko | "Deep Creek Reservoir, along the IL Ranch Road, 10.5 km west of the main NS road." | 7/14/02 |
| 97 | Marsila vestita | A. Tiehm 14569 | UTC | 244995 | NV: Elko | "Owyhee Desert, Josephine Reservoir, 2.4 road miles NW ot Butte Springs." | 6/26/04 |
| 98 | Marsilea vestita | R.J. Shaw 3708 | UTC | 187738 | UT: Millard | North of Flowell. | 9/9/84 |
| 99 | Marsilea vestita | Weber & Wittmann 18558 | UTC | 212240 | CO: Baca | "Big Hole Canyon, above Est fort Carrizo Creek, Road H" | 5/22/93 |
| 100 | Marsilea vestita | R.J. & M. Shaw 4986 | UTC | 208944 | MT: Lake | "Ninepipes Reservoir Wildlife Refuge, near Charlo; 150 yards above dam." | 9/11/91 |
| 101 | Marsilea minuta | A.R. Diamond 14269 | MISS | 70573 | AL: Pike | "Conecuh River, US Hwy 29, east side of river, north of the road." | 11/16/03 |
| 102 | Marsila minuta | D.M. Johnson 800 | MICH | Trinidad: Nariva Cocal | "Nariva Cocal, 5 km S of point where Manzanilla-Mayaro Road reaches Manzanilla Bay." | 3/18/82 | |
| 103 | Marsilea deflexa | D.M. Johnson 794 | MICH | Venezuela: Guarico | "Km 239 on highway from San Fernando de Apure to Calabozo, ca. 36 km N of Camaguan." | 3/13/82 | |
| 104 | Marsilea ancylopoda | D.M. Johnson 773 | MICH | Argentina: Corrientes | "Depto. Mburucuya, Estancia Sanda Maria" | 2/12/82 | |
| 105 | Marsilea ancylopoda | E. Lott & A. Sanders 3987 | MICH | Mexico: Jalisco: Quemaro | "Mpio. La Huerta; Quemaro, ca. 25 km nw of Esatcion de Biologia Chamela; pond on road to Las Alamandas." | 11/2/91 | |
| 106 | Marsilea minuta | M. Dyer 173 | MICH | Nigeria: Kano: Acha Lafia | Acha Lafia. | 2/2/80 | |
| 107 | Marsilea polycarpa | W. Wagner | MICH | Puerto Rico: Loiza | "Loiza, Pinones Munic., grazed island in white and black mangroves." | 9/23/82 | |
| 108 | Marsilea polycarpa | D.M. Johnson 793 | MICH | Venezuela: Apure | "Pond along road from Mantecal to Elorza, ca. 25 km N of Elorza." | 3/12/82 | |
| 109 | Marsilea mollis | N. Murray & D.M. Johnson 1404 | MICH | Mexico: Chiapas | Km 152 on S side of Hwy 190 just W of San Francisco between Laguna Larga and Ajasaxh. | 6/3/83 | |
| 110 | Marsilea vestita | B. Erttter et al. 8131 | MICH | CA: Contra Costa | "Alkali sink W and adjacent to Clifton Court Forebay, ca. 15 air miles SW of Stockton." | 4/5/89 | |
| 111 | Marsilea macropoda | D.M. Johnson 721 | MICH | TX: Aransas | Along Farm Road 1781 just north of Aransas Co. airport | 5/23/81 | |
| 112 | Marsilea coromandelina | M. Dyer 172 | MICH | Nigeria: Kano: Acha Lafia | Acha Lafia | 2/2/80 | |
| 116 | Marsilea mutica | Page s.n. | VPI | VA: Hanover |